


Glucagon signaling pathway overview
The glucagon signaling pathway refers to the sum of a series of proteins and regulatory factors involved in the function of glucagon. Human pancreatic hyperglycemia is a linear polypeptide consisting of 29 amino acids with a molecular weight of 3485, which is also cleaved by precursors of macromolecules. The concentration of pancreatic hyperglycemia in serum is 50-100 ng/L, and the half-life of plasma is 5-10 min. In contrast to the role of the insulin signaling pathway, the glucagon signaling pathway is a pathway that promotes catabolism. The glucagon signaling pathway has a strong role in promoting glycogenolysis and gluconeogenesis, resulting in a significant increase in blood glucose. 1 mol/L hormone can rapidly decompose 3 x 106 mol/L glucose in glycogen. The glucagon signaling pathway activates hepatocyte phosphorylase and accelerates glycogenolysis through the cAMP-PK system. The gluconeogenesis is enhanced as hormones accelerate the entry of amino acids into the liver cells and activate the enzyme system involved in the gluconeogenesis process. The glucagon signaling pathway also activates lipase, which promotes fat breakdown, while at the same time enhancing fatty acid oxidation and increasing ketone body formation. The target organ of the glucagon signaling pathway that produces the above metabolic effects is the liver, which removes the liver or blocks the blood flow of the liver (Figure 2), and these effects disappear. In addition, the glucagon signaling pathway promotes the secretion of insulin and islet somatostatin. Pharmacological doses of glucagon can increase cAMp content in cardiomyocytes and increase myocardial contraction.
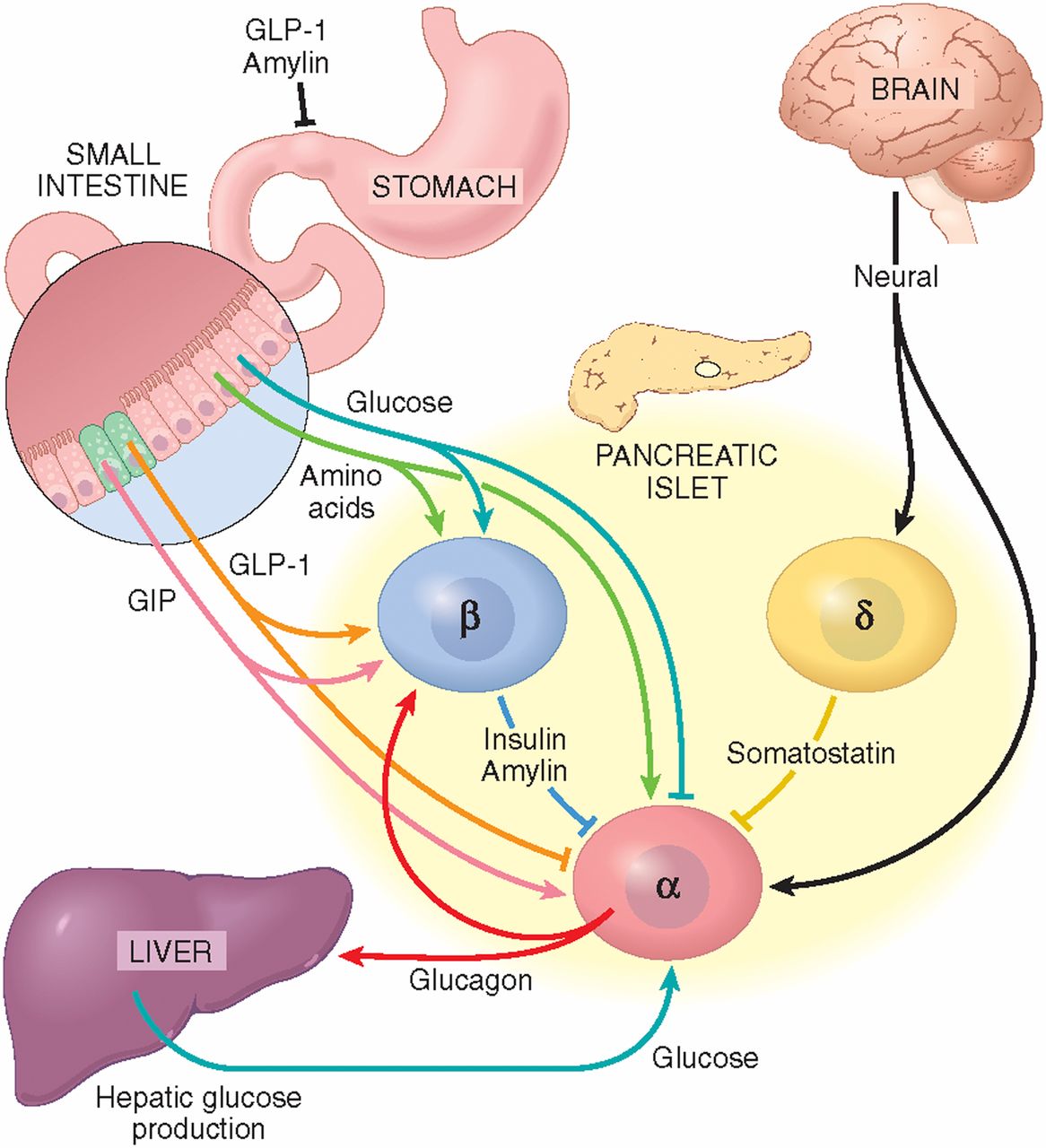
Figure 2. Individual controls of glucagon secretion
Glucagon signaling pathway family
The glucagon receptor belongs to a 4B family of receptors formed by seven transmembrane G protein couplings. It is mainly distributed in the liver, and followed by tissue cells such as kidney, muscle, fat, brain, intestine, adrenal gland, spleen, ovary, thyroid gland, and pancreatic islet(α and β cells). Such receptors are characterized by being found located on the cell surface and conjugated to the G protein. When glucagon secreted by islet alpha cells binds to the glucagon receptor on the surface of the target tissue cells, the glucagon receptor conformation changes and the G protein is activated. There are many types of G proteins, of which Gsα and Gq are related to the glucagon receptor. When Gsα is activated, adenylate cyclase is activated and intracellular cAMP production is increased, which in turn activates protein kinase A (PKA), and leads to phosphorylation of functional proteins in the cells. This pathway is called PKA pathway. When Gq is activated, the phospholipase C-inositol triphosphate channel (PLC-INSP3) pathway can be activated, causing intracellular Ca2+ release. The above two pathways will directly or indirectly cause a decrease in glycolysis, a decrease in glycogen synthesis, an increase in gluconeogenesis, an increase in glycogenolysis, and eventually an increase in blood glucose. Under physiological conditions, insulin secreted by β cells inhibits α by paracrine action. The cells secrete glucagon; on the other hand, glucose also inhibits glucagon secretion. In pathological conditions, such as diabetes, this negative feedback balance is disrupted due to impaired insulin secretion or insulin resistance in alpha cells, and glucagon levels are significantly elevated. Unger discovered in 1978 that elevated blood glucose in type 2 diabetic patients did not normally inhibit glucagon secretion. Larsson and Ahren used the venous amino acid stimulation test and the oral glucose tolerance test in a population with impaired glucose tolerance (IGT), respectively, and found that there was inappropriate hyperglycemic secretion after a meal and could not be inhibited by insulin. Therefore, the normality of the insulin signaling pathway requires all members of this pathway to work together.
Glucagon signaling pathway
- Glucagon signaling cascade
- The glucagon signaling pathway is initiated by binding to specific receptors on the target cell membrane, which activates adenylate cyclase by Gs protein, catalyzing the conversion of adenosine triphosphate (ATP) to cyclic adenosine monophosphate (cAMP), thereby increasing intracellular cAMP levels. cAMP is the major second messenger of glucagon glycosylation, exerting excitatory effects on pancreatic alpha cells secreting glucagon by two pathways (cAMP-dependent protein kinase A pathway and non-cAMP-dependent protein kinase pathway). Alpha cells secrete glucagon through different ion channels, and the ATP-sensitive K channel is considered to be the main channel, mainly regulating glucagon secretion.
- Pathway regulation
- In recent years, researchers have intervened in the glucagon signaling pathway through various methods, including intervention of pre-receptor (pre-binding) regulation, regulation of receptor binding, and post-receptor (post-binding) regulation to achieve lowering of blood glucose and treatment of diabetes. Re-receptor intervention method: by improving insulin resistance in islet alpha cells and reducing glucagon secretion. Weiss et al found that the conversion of NGT to IGT was accompanied by a decrease in insulin sensitivity, accompanied by a gradual increase in glucagon secretion. The expression is up-regulated and insulin resistance is maintained in islet alpha cells. As mentioned above, when the alpha cell insulin is resistant, its signal transduction pathway is impaired. Exploring its mechanisms may be related to the mediation of inflammatory mediators. Studies have shown that inflammatory factors play an important role in peripheral insulin resistance, and the effect of nuclear factor kappa B (NF-κB) on alpha cells in a model of insulin resistance in rat islet alpha cells induced by high-fat feeding mediates activation of the inflammatory pathway. Ellingsgaard et al found that IL-7 receptors were expressed on islet α cells compared with other tissues. IL6 induced the expression and secretion of glucagon in rats with high-fat diet. After using the IL6 receptor gene knockout model, the body's metabolic disorder was corrected. The use of thiazolidinediones (TZD) drugs can not only improve peripheral insulin resistance in SD rats induced by high-fat feeding, but also inhibit the proliferation of α cells, and and significantly increase glucagon levels and α-cell glucagon mRNA expression. This effect is achieved by the binding of TZDs to the peroxisome proliferator-activated receptor on islet alpha cells, which directly inhibits glucagon gene transcription. In recent years, there are many studies on the treatment of diabetes with incretin hormone, which is represented by glucagon like peptide1 (GLP1) and its analogs. GLP1 is a 30 amino acid peptide hormone secreted mainly by L cells of the distal ileum, rectum and colon. It not only acts on glucose-dependent β-cells, but also promotes insulin secretion. It also acts on islet α cells. Inhibition of glucagon secretion can improve alpha cell insulin resistance. Prohormone converting enzyme 2 (PC2) gene knockout: proglucagon is a precursor of glucagon, which produces different products through different prohormone convertases in different tissue organs. Among them, glucagon is produced by PC2 in the pancreas, and GLP1 and GLP2 are produced in the intestine via PC1/3. Study have showed that PC2 knockout mice have a significant decrease in blood glucagon, mild persistent hypoglycemia, and modern compensatory islet alpha cell proliferation, when using a micro-osmotic pump or intraperitoneal small dose. After glucagon injection, blood glucose returned to normal; and after a long period of application, the morphology of islet α cells recovered to resemble that of wild-type mice. Glucagon neutralizing antibodies: this method uses exogenous glucagon antibodies to bind to glucagon in the body, thereby blocking the effects of endogenous glucagon and ultimately lowering blood sugar. The brand is equivalent to an experiment conducted in 1996 using a high-capacity, high-affinity glucagon monoclonal antibody (Glu-mAb) in a normal, alloxan (ALX)-induced mild and severe diabetic rabbit model. Tip: this antibody can completely block exogenous glucagon-induced hyperglycemia in normal animals; in low-glycemic zoos, lowering blood sugar is also obvious; in high-glycemic type 1 diabetic rabbits, Glu-mAb can still significantly reduce liver glucose output, reducing the fasting blood glucose of experimental rabbits (from 21.2 mmol / l to 12. 7mmol / l). The use of glucagon antibodies to reduce the effects of glucagon can better control the effects of type 2 diabetes. Barbato et al. found that the 40-glycine-serine polymorphism (Gly40Ser) of the glucagon receptor gene exon 2 in French Caucasians is closely related to type 2 diabetes. The research focused on glucagon receptor blockers, glucagon receptor gene expression inhibitors, and glucagon receptor gene knockout. Receptor blockers: the mechanism of action of glucagon receptor blockers is mainly through competitive binding to endogenous glucagon, thereby inhibiting glucagon-mediated adenylate cyclase activity, reducing glycogen output, reducing fasting blood glucose levels, and improving glucose tolerance. The receptor blocker is classified into a peptide compound and a non-peptide small molecule compound according to the molecular structure. Petersen et al. found that a non-peptide small molecule compound, Bay 27 9955, effectively blocks the increase in glucose production and blood glucose caused by exogenous glucagon in healthy adult males. This is also the only drug that has been used in humans for glucagon receptor antagonists. Although more clinical trials are needed to prove efficacy, it is undoubtedly an increase in the search for effective human glucagon receptor antagonists. The above studies have shown that both glucagon receptor antagonists, whether peptide or non-peptide, block the liver glucagon receptor and exert a hypoglycemic effect. Receptor gene expression inhibitors: the principle of action of these drugs is to block the expression of glucagon target receptor gene and reduce the expression of glucagon receptor mRNA, thereby achieving the role of treating diabetes. Sloop and other antisense oligonucleotides (ASO) blocking glucagon receptors were used to treat type 2 diabetic animals. It was found that glucagon receptor mRNA expression decreased and plasma glucagon concentration increased significantly after treatment. Glucose tolerance improved, and triglycerides and free fatty acids decreased significantly. Post-receptor regulation: there are still few intervention studies on glucagon receptors, but there are still some reports on G-protein coupled receptor alpha knockout animals. G protein-coupled receptors are present in multiple organs throughout the body. The glucagon receptor is mainly in the liver. The use of liver-specific G protein knockout animals is a method of interfering with the glucagon signaling pathway.
- Relationship with diseases
- Type II diabetes
- Activation of glucagon signaling pathways and dysfunction play an important role in the pathophysiology of type 2 diabetes. More and more studies have shown that the decrease of the secretion of glucagon by inhibiting alpha-cell production, neutralizing blood circulation and hyperglycemia, changing glucagon receptor gene expression, and other methods to interfere signaling pathways may be new treatments for diabetes.
- Obesity
- Recent studies have shown that obese patients have both dysfunction of islet β-cells and α-cells, impaired insulin secretion and excessive secretion of glucagon, which aggravates the disorder of blood glucose metabolism, so the glucagon signaling pathway regulated for obese patients treatment is especially important.
- Coronary heart disease
- In the past two years, the levels of insulin and glucagon in the patients with coronary heart disease were significantly higher than those in the control group, and the inhibitors of the glucagon signaling pathway were improved, so the glucogon signaling pathway was involved in coronary heart disease. But the detail has to be further studied.
References:
- Charron, M, and P. Vuguin. Lack of glucagon receptor signaling and its implications beyond glucose homeostasis. Journal of Endocrinology. 2015, 224(3):R123
- Hamilton A, Zhang Q, Salehi A, et al. Adrenaline Stimulates Glucagon Secretion by Tpc2-Dependent Ca2+ Mobilization from Acidic Stores in Pancreatic α-Cells. Diabetes. 2018:db171102.
- Cheng X, Kim S Y, Okamoto H, et al. Glucagon contributes to liver zonation. Proceedings of the National Academy of Sciences. 2018:201721403.
- Lapierre M P, Abraham M A, Filippi B M, et al. Glucagon and lipid signaling in the hypothalamus. Mammalian Genome. 2014, 25(9-10):434-441.
- Rah S Y, Kim U H. CD38-mediated Ca(2+) signaling contributes to glucagon-induced hepatic gluconeogenesis. Scientific Reports. 2015, 5:10741.
Comments